")
")






 Otx-гены — древнее семейство гомеозисных генов, которые действуют по тому же принципу, что и широко известные Hox-гены, но не в туловище, а в передней части головы. Работа этих генов очень важна для развития передних отделов мозга позвоночных, и в частности — коры наших больших полушарий. Итальянские ученые показали, что мутации по этим генам, изменяя распределение нейронов в коре, могут приводить к эпилепсии, но могут и защищать от нее. При этом Otx-гены есть у насекомых, у червей и даже у кишечнополостных. Их новые функции, связанные с регуляцией развития коры больших полушарий, безусловно, появились вместе с самой корой, то есть только у высших позвоночных.
Otx-гены — древнее семейство гомеозисных генов, которые действуют по тому же принципу, что и широко известные Hox-гены, но не в туловище, а в передней части головы. Работа этих генов очень важна для развития передних отделов мозга позвоночных, и в частности — коры наших больших полушарий. Итальянские ученые показали, что мутации по этим генам, изменяя распределение нейронов в коре, могут приводить к эпилепсии, но могут и защищать от нее. При этом Otx-гены есть у насекомых, у червей и даже у кишечнополостных. Их новые функции, связанные с регуляцией развития коры больших полушарий, безусловно, появились вместе с самой корой, то есть только у высших позвоночных.
Хорошо известно, что в геномы всех современных многоклеточных организмов входит множество очень древних генов. По данным сравнительной геномики, характерное время жизни генов животных измеряется сотнями миллионов лет (Е. В. Кунин. Логика случая. М., 2014). В нашем геноме есть и гены, возраст которых превышает миллиард лет (см.: У одноклеточных организмов есть ген, способный управлять развитием хорды, «Элементы», 25.10.2013). Это неудивительно, если учитывать, что около 80% генов человека являются общими как минимум для всех позвоночных.
Важное место среди древних генов занимают гены, контролирующие развитие целых отделов тела. Они называются гомеозисными генами. Особенно широко среди них известны гены семейства Hox, регулирующие у зародышей становление сегментации (см., например: Программы работы Hox-генов у личинок и взрослых особей кольчатого червя принципиально отличаются, «Элементы», 27.05.2013). Hox-гены есть у всех двусторонне-симметричных животных, а также у кишечнополостных. Это означает, что, к примеру, у общего предка человека, осьминога и медузы хотя бы один Hox-ген уже точно был.
Еще одно важное семейство гомеозисных генов называется Otx. Гены Otx — в некотором смысле антагонисты генов Hox. Дело в том, что зона активности (правильнее говоря, экспрессии) генов Hox никогда не охватывает всего тела. В передней области головы — там, где находятся глаза, передние отделы мозга, усики насекомых или челюсти позвоночных, — так вот, в этой части тела животных экспрессии Hox-генов нет. Зато там обязательно экспрессируются Otx-гены (рис. 1). Можно сказать, что они работают «вместо» Hox-генов, вызывая развитие совсем иных органов, чем в туловище.
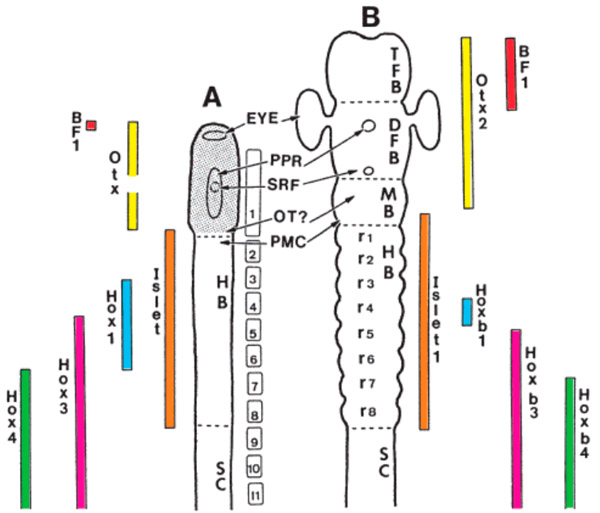
Рис. 1. Экспрессия гомеозисных генов в передней части центральной нервной системы у ланцетника (A) и позвоночного (B). TFB — передний мозг, DFB — промежуточный мозг, MB — средний мозг, HB — продолговатый мозг, SC — спинной мозг, EYE — глаз; остальные анатомические детали в тексте не упоминаются. Разноцветные столбики показывают протяженность областей экспрессии вдоль нервной трубки. Например, голубой столбик обозначает область экспрессии гена Hox1, фиолетовый — гена Hox3, зеленый — гена Hox4, а желтый — интересующего нас сейчас в первую очередь гена Otx2. У ланцетника соответствующий ген называется просто Otx, потому что у него он один
Otx-гены есть у всех двусторонне-симметричных животных (рис. 2), и у кишечнополостных тоже. Вероятно, они возникли примерно тогда же, когда и Hox-гены, то есть более 500 миллионов лет назад.
|

У млекопитающих имеется два Otx-гена, которые так и называются Otx1 и Otx2. Оба они участвуют в развитии центральной нервной системы. Область экспрессии Otx2 тянется через весь передний, промежуточный и средний мозг (рис. 1, B). Область экспрессии Otx1 чуть меньше; она охватывает те же отделы мозга, но находится целиком внутри области экспрессии Otx2.
Группа итальянских исследователей заинтересовалась вопросом: а не связаны ли повреждения генов Otx с заболеваемостью эпилепсией? Вопрос этот вполне оправдан. Эпилептический приступ представляет собой вспышку сильного возбуждения, обычно возникающую в коре больших полушарий. А кора больших полушарий — это часть переднего мозга (telencephalon), того самого отдела ЦНС, для развития которого Otx-гены особенно важны.
Объектами опытов итальянцев стали мыши, а подход был применен самый классический: «сломать и посмотреть, что получится»; благо, генная инженерия давно уже позволяет выключить (нокаутировать) какой угодно ген. И сразу выяснилось, что выключение гена Otx1, в полном соответствии с ожиданием, порождает самых настоящих мышей-эпилептиков. Они то навязчиво дергают головой, то крутятся на месте, то впадают в конвульсии, и электроэнцефалография при этом показывает картину, типичную для эпилептических приступов.
Детальное исследование показало, что в мозге мышей, нокаутных по гену Otx1, наблюдается целый ряд морфологических отличий от нормы (рис. 3, A, B).
Во-первых, у них оказались уменьшены общая толщина коры больших полушарий и размер гиппокампа — эволюционно древнего участка коры, работа которого связана с эмоциями и с памятью.
Во-вторых, нарушено распределение по коре так называемых пирамидных клеток, или пирамидных нейронов. Это основные эфферентные нейроны нашей коры, передающие возбуждение из нее на другие структуры. Давно установлено, что кора больших полушарий мозга млекопитающих состоит из шести клеточных слоев, которые принято обозначать римскими цифрами. Пирамидных клеток в норме больше всего в V слое, который иногда так прямо и называют пирамидным. У мышей с выключенным геном Otx1 в V слое коры значительно меньше пирамидных нейронов, чем положено, а во II и III слоях, наоборот, значительно больше.
В-третьих, выключение гена Otx1 нарушает расположение клеток, имеющих длинное название парвальбумин-позитивные ГАМК-эргические интернейроны. Разобраться в смысле этого названия не так уж сложно. Парвальбумин — это кальций-связывающий белок, содержание которого удачно маркирует некоторые клеточные популяции. Интернейроны, или вставочные нейроны, — это нервные клетки, передающие сигналы с одних нервных клеток на другие (а не на мышцы, например). И наконец, ГАМК, или гамма-аминомасляная кислота, — это медиатор, то есть выделяемое нейронами вещество, действующее на другие нейроны. Особенность ГАМК по сравнению с другими медиаторами в том, что ее эффект — тормозящий: она не усиливает возбуждение клеток, на которые действует, а гасит его. ГАМК-эргические нейроны — это тормозные нейроны. У мышей, нокаутных по гену Otx1, распределение ГАМК-эргических нейронов в коре больших полушарий становится резко неравномерным, пятнистым («где густо, где пусто»), и это означает, что там нарушена система торможения.
Таким образом, связь гена Otx1 с эпилепсией прояснена. Ведь эпилептический приступ — это, по сути, неконтролируемое растекание возбуждения по коре. Естественно, что при нарушении баланса возбуждения и торможения, и особенно при ослаблении самого торможения, наступление таких приступов облегчается.
В высшей степени показательно, что если вместо «сломанного» гена Otx1 вставить в мышиный геном гомолог этого гена, взятый у мухи-дрозофилы (он называется otd), то все вредные последствия исчезают. Продукты Otx-генов позвоночных и членистоногих, общий предок которых, скорее всего, жил еще в докембрии, до сих пор являются вполне взаимозаменяемыми. Такая консервативность не уникальна, она вообще характерна для гомеозисных генов, но всё равно этот результат впечатляет.
С геном Otx2 дело обстоит чуть сложнее. Полное выключение этого гена ни к каким интересным результатам не приводит, потому что оно летально: у таких зародышей просто отсутствует вся передняя часть мозга и они погибают на ранних стадиях. Исследовать там нечего. Но существуют мутации, которые не выключают этот ген полностью, а или слегка ослабляют, или слегка усиливают его активность. Эффекты этих мутаций тоже были изучены (рис. 3, C, D). Оказалось, что они влияют в первую очередь на средний мозг, а конкретно — на те нейроны среднего мозга, которые выделяют медиатор дофамин. При ослабленной активности гена Otx2 (рис. 3, C) дофаминэргических нейронов в среднем мозгу становится меньше, чем в норме; их предшественники дифференцируются по другому пути и начинают вместо дофамина выделять серотонин. При повышенной активности гена Otx2 (рис. 3, D) дофаминэргических нейронов в среднем мозгу, наоборот, больше, чем в норме. Тут эффект оказывается многоступенчатым. Отростки дофаминэргических нейронов среднего мозга тянутся в передний мозг и там, видимо, как-то влияют на дифференцировку тех самых ГАМК-эргических нейронов, о которых мы уже говорили в связи с геном Otx1. Чем больше дофамина, тем ГАМК-эргических нейронов в коре переднего мозга больше. А это, в свою очередь, должно влиять на вероятность наступления эпилепсии. ГАМК-эргические нейроны — тормозные, они должны гасить эпилептический приступ; и действительно, у мутантов с их повышенным числом устойчивость к таким приступам выше.
Самое интересное, что и у мутантов со сниженной активностью гена Otx2 вероятность эпилепсии не повышается. Наоборот, устойчивость к некоторым типам приступов у них даже возрастает. Связано это с тем, что у таких мутантов в среднем мозге увеличивается число серотонинэргических нейронов, отростки которых тоже идут в кору больших полушарий. А серотонин в центральной нервной системе часто действует как тормозной медиатор, и его эффект — антиэпилептический.
Таким образом, если выключение гена Otx1 провоцирует эпилепсию, то мутации по гену Otx2 от нее скорее защищают. Очень примечательно, что последнее касается разных мутаций — как ослабляющих, так и усиливающих действие этого гена. Это интересный пример ситуации, когда противоположно направленные воздействия дают одинаковый конечный эффект.
|

Всё это, разумеется, не значит, что Otx-гены являются какими-то особыми «генами эпилепсии». Эпилепсия — очень неспецифическая вещь, и приводить к ней, и защищать от нее могут десятки разных факторов. Просто мы теперь знаем, что мутации Otx-генов в число таких факторов несомненно входят.
Одновременно вся эта история дает нам очередной повод задуматься о путях эволюции. Сравнительная геномика показала нам, что гены — вещь необычайно стабильная. Новые гены появляются в эволюции многоклеточных животных явно реже, чем возникают новые органы. Но ведь развитие новых органов тоже надо как-то регулировать, и поэтому древние гены просто вынуждены менять функции.
Вот мы теперь узнали, что у млекопитающих ген Otx регулирует распределение нейронов по слоям коры больших полушарий. Но у большинства животных, имеющих этот ген, никакой коры и никаких полушарий вообще нет! Это хороший пример того, насколько легко древний ген может приобрести функцию, немыслимую раньше. Можно сказать, что геном одновременно очень консервативен и очень пластичен: консервативен по части состава генов и пластичен в области их функций, которые постоянно обновляются.
Выдающийся генетик Н. В. Тимофеев-Ресовский однажды сказал: «У человека не может возникнуть мутации, изменяющей величину и форму кисточки на конце хвоста, потому что у Homo sapiens к этому нет онтогенетических предпосылок» (Н. В. Тимофеев-Ресовский, Н. Н. Воронцов, А. В. Яблоков. Краткий очерк теории эволюции. М., 1977). Развивая эту мысль, мы вправе добавить: но вполне вероятно, что ген, потенциально способный менять величину и форму кисточки на хвосте, у человека при этом все-таки есть! Ген — это почти универсальный управляющий элемент, средой для работы которого является развитие организма (онтогенез). Вполне естественно, что управлять он может разными процессами, — смотря чего конкретный организм потребует. В этом отношении Тимофеев-Ресовский был совершенно прав.





Рис. 2. Экспрессия некоторых гомеозисных генов в зародышах мухи-дрозофилы (A) и лягушки (B). Заштрихованная область — закладка центральной нервной системы; у мухи она находится на брюшной стороне, у мыши на спинной. Стрелками указана область экспрессии гена, маркирующего ту сторону тела, где центральной нервной системы нет; у насекомых этот ген называется dpp, а у позвоночных BMP-4. А поперечная пунктирная линия делит центральную нервную систему на области экспрессии Hox (задняя часть головы и туловище) и Otx/otd (передняя часть головы). Это деление у позвоночных и членистоногих выглядит сходно. Не вызывает сомнений, что ген, который у позвоночных называется Otx, а у насекомых otd, уже был у их общего предка